Mechanisms of cell and tissue polarity formation
Membrane proteins are mobile in the lipid fluid environment. In contrast, PIN proteins are localized at specific regions of plasma membranes (PMs). What mechanisms can determine the polar localization of PIN proteins? It has been widely believed that the membrane trafficking system, which involves recycling of PIN proteins from endosomes to a specific region of the PM, determines the polar localization of PIN proteins. However, we take respectful issue with this hypothesis. Instead, we demonstrated that PIN proteins are organized into a cluster-like structure, whose mobility and endocytosis at the PM are inhibited. Furthermore, we found that PIN clusters are biomolecular condensates involved in maintaining polar localization and regulating efflux activity of PIN proteins. We are currently studying the molecular mechanism of PIN cluster formation to elucidate the mechanisms of the polar localization of PIN proteins. This study will be carried out from the perspective of membrane dynamics, membrane trafficking and cytoskeleton. The physiological function of PIN clusters, including the venation pattern and the tropism (Phototropism, Gravitropism and Halotropism), will also be investigated.
Current projects related to the mechanisms of cell and tissue polarity formation are:
1. Cell polarity and tissue patterning by auxin transporter PIN proteins
- Regulatory mechanisms of PIN cluster formation and reestablishment
- Modeling of auxin canalization hypothesis
2. Cell polarity formation mediated by membrane trafficking and cytoskeleton dynamics
- Control of cell membrane properties by VAN3 ARF-GAP
- Phospholipid metabolism in plasma membrane
- Crosstalk of small GTPases at the cell surface
Learn more about the research
1. Cell polarity and tissue patterning by auxin transporter PIN proteins
Regulatory mechanisms of PIN cluster formation and reestablishment
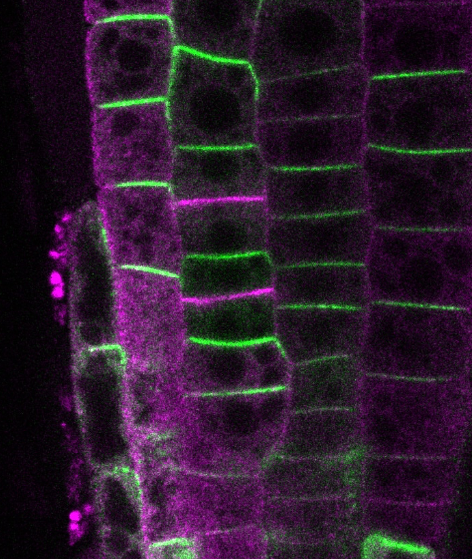
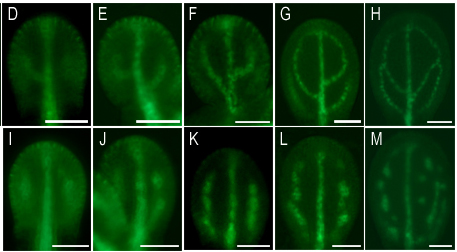
多細胞生物の発生においては、細胞極性と体軸が適切に形成されることが必要です。維管束植物の体軸は、オーキシンの極性輸送により形成されますが、その輸送方向はオーキシン担体PINの細胞における極性局在(細胞膜の特定部位への偏在)によって規定されています。近年、わたしたちはPINの局在制御を担う新しいメカニズムについて重要な発見をしました。PINが細胞膜中で形成する多量体構造「PINクラスター」が、PINの膜中の流動性を抑制する機能を発揮し、極性局在の確立に寄与するというものです。
わたしたちの研究室では、現在、クラスターの分子構造、化学的性質、形成メカニズムについて研究を行っています。これまでにわたしたちは、PINクラスターの形成において働く主要なメカニズムを同定することに成功しました。また、PINクラスターの形成は、PINの細胞膜上での局在の安定化だけでなく、オーキシンの排出活性の制御などに働くことを示唆する結果も得ました。今後、これらの作業仮説の実証を行うとともに、PINの局在性御に関わる既知の因子とPINクラスターの関係や、PINの局在性御に関わる新奇因子の解析を行うことで、オーキシン極性輸送メカニズムの解明を目指します。
Modeling of auxin canalization hypothesis
これまでの生理学実験から、オーキシンは自律的に細胞間を流れ続ける性質があるとの仮説が提唱されており、オーキシンキャナライゼーション仮説と呼ばれています。オーキシンには、細胞におけるオーキシンの排出活性を正に制御する作用があり、この正のフィードバック作用により、オーキシンは自律的に流路を形成すると想定されています。わたくしたちは、現在、PINクラスターの発生過程における挙動について詳細に研究を行っています。これまでに、PINクラスターは細胞分裂過程で特徴的な挙動を示すことを見出しました。今後、細胞内のオーキシン動態の可視化や、細胞レベルでのオーキシンの摂動実験、さらには数理モデルなどの実験にも挑戦することで、オーキシンキャナライゼーション仮説の検証や、細胞極性と体軸の形成について新しいモデル(仮説)の構築・検証を行います。
2. Cell polarity formation mediated by membrane trafficking and cytoskeleton dynamics
Control of cell membrane properties by VAN3 ARF-GAP
現代の生物学では、遺伝子やタンパク質の機能についての知見が蓄積された結果、細胞内の現象を、個々の分子の機能だけにとどまらず、複数の分子集合体が織りなすダイナミズムとして理解することができるようになってきました。
PINの局在制御についても同様で、PINが小胞輸送されて細胞膜に運ばれる、といったレベルの理解ではなく、「PIN、PINクラスター構成分子、PIN・PINクラスター構成因子と相互作用しクラスター形成のON/OFFを制御する分子、細胞膜のリン脂質など、が複合的に関係しあって、PINが極性局在する場を形成する」という世界観で研究することが可能となってきています。
わたしたちが研究している分子には、このような世界観を広げてくれる候補分子が複数あります。とくに、VAN3はBARドメイン、PHドメインという細胞膜、脂質に関連したドメインを持つことがわかっていて、このことから多様な機能を持つ可能性が感じられます。まず、BARドメインは、生体膜の湾曲度合いの認識や、膜自体を湾曲させる機能を持つと言われています。また、細胞骨格(アクチン、微小管)と結合して機能することが知られています。そこで、VAN3のBARドメインが膜の湾曲へ関与しているのか?細胞骨格と連携して機能しているかについて解析を行います。VAN3のBARドメインと生体膜の関係については生化学解析、VAN3と細胞骨格との関係についてはイメージングなどの手法を用いて研究していきます。
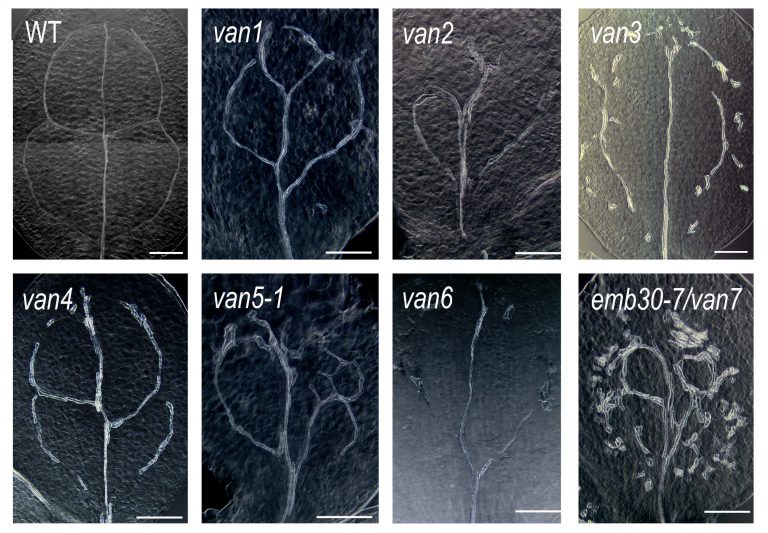
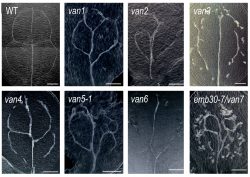
(Koizumi et al., 2000)
Phospholipid metabolism in plasma membrane
VAN3がもつPHドメインは、リン脂質に結合します。実際、わたしたちは、VAN3の局在と活性は生体膜のリン脂質環境によって制御されることを明らかにしました(Naramoto et al., 2009 Development)。また、この制御を担っている因子も複数同定しています。
そのなかのひとつでリン脂質合成酵素のCVP2(cotyledon vascular pattern2) は、細胞の頂端部に限定的に局在するという独自の局在パターンを示します。また、葉脈以外のパターン形成との関係を示唆する結果も得られており、注目しています。今後、CVP2がどのような発生学的機能を果たしうるのか、分子・細胞・個体レベルで解析を行います。
Crosstalk of small GTPases at the cell surface
VAN3は、どのようなタンパク質と相互作用するかが既に調べられています。そのひとつのVAB(VAN3 binding protein)は詳しい機能は分かっていませんが、生体膜のリン脂質と結合します(Naramoto et al., 2009 Development)。また、細胞内の生命現象に重要な役割を果たすsmall GTPase スーパーファミリーであるRabやRopと相互作用する可能性も示唆されています(楢本、未発表)。
RopやRabは分子スイッチとして多種多様な現象に関わっています。動物では、 ARHGAPやARAPというARFとRhoの両者と相互作用するドメインを持つ分子がありますが、植物にはありません。またVABは植物特異的な分子です。VAN3がVABを介してRopとRabとどのように関係し合っているのかが明らかになれば、small GTPaseが他の因子とクロストークする仕組みについて全く新しい知見が得られると期待しています。