常に世界にさらされながら、体のかたちを作っていく仕組み
植物は動きません。生えている場所で環境変化に見舞われてしまったとき、植物はどのように対応しているのでしょうか?植物の環境応答の仕組みは、形態形成のときに使われる仕組みと深いつながりがあることがわかっています。
現在取り組んでいる課題、これから取り組みたいと考えている課題は次のとおりです。
1. 不等分裂の研究:コケ胞子の第一分裂で生じる非対称性とは?
2. 光・重力に依存した植物の発生制御機構の解析:コケの幹細胞の位置が光と重力シグナルによって決まる仕組みとは?
3. 環境応答時のPINクラスターとVAN3を中心とした膜ダイナミクスの動態についての研究
4. 環境ストレス(土壌環境・病原菌応答)応答における細胞膜ドメイン動態
研究内容をさらに詳しく
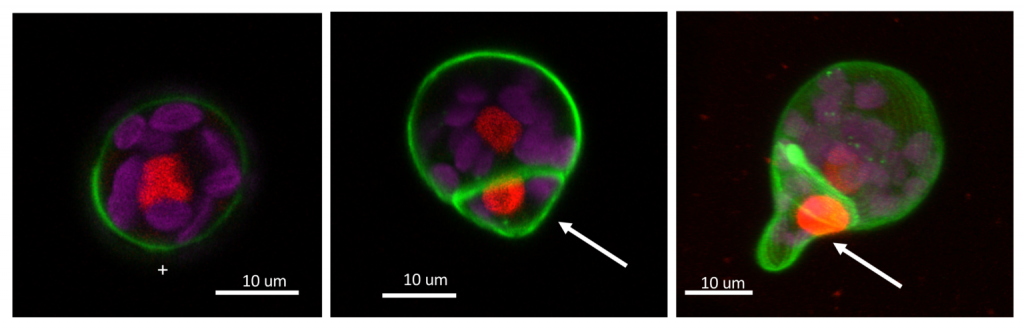
1. 不等分裂で生じる非対称性とは?
形態や運命が異なる娘細胞が生み出される分裂は、不等分裂と呼ばれ、多細胞生物では発生のさまざまなステージで観察されます。とりわけ、細胞壁があり細胞が移動できない制約をもつ植物においては、発生・形態形態においては、生み出された細胞は隣接した細胞との位置関係が全く変わらないまま、適切な分化と成長を遂げる必要があります。こういうときに、不等分裂が重要な役割
を果たします。具体的には、胚発生における初期の分裂は動物、植物ともに不等分裂がおきます。また、植物の幹細胞周辺で恒常的に起きる細胞分裂も不等分裂です。不等分裂では一般に、タンパク質やmRNA等の存在量・分布の偏り、細胞内の膜構造、細胞骨格の動的な変化が生じることが報告されています。植物の世界でもたくさんのバリエーションの不等分裂を見出すことができますが、わたしたちはコケ植物の胞子の第一分裂に注目しています。
コケ植物の第一分裂で生み出される娘細胞は、一方は将来幹細胞になる未分化な細胞、もう一方はそれ以上分裂・分化しない仮根に分化した細胞となります。類似の不等分裂は、褐藻の受精卵等においても観察され、植物の広い種で保存されています。この不等分裂で生じる非対称性の実態などを明らかにしていきたいと考えています。
わたしたちは近年、コケ植物の胞子は、光や重力に応じて、分裂面が制御されることを示唆する結果も得ました。今後、ゼニゴケ胞子の不等分裂メカニズムを理解することで、生物の発生・形態形成の基盤となる細胞の不等分裂機構の理解を目指します。
2. 光・重力に依存した植物の発生制御機構の解析:コケの幹細胞の未分化性が光と重力シグナルによって制御される仕組みとは?
ゼニゴケをオーキシン極性輸送阻害剤の存在下で生育させると、背腹軸が乱される特徴的な表現型があらわれます(楢本、未発表)。また、類似の表現型は、生育の際に照射する光の方向をランダムにした場合にも観察されます。これらの表現型は、クリノスタットを用いた微小重力条件(無重力を模した環境)で生育させた際に生じる表現型として過去に報告されているものと類似しています。具体的には、ウルトラセブンに登場する双頭の怪獣パンドンを連想させるので、この表現型を「パンドン型」と呼んでいます(検索してみてください)。パンドン型の表現型は、pin1変異体、PINの局在制御因子VAN3、活性制御因子D6PKのゼニゴケホモログの変異体においても観察されます(楢本、未発表)。このことから、背腹軸の形成はオーキシンとPINの機能に依存的であり、PINの制御因子群もこのプロセスで機能すると考えられます。
現在、わたしたちは、パンドン型の表現型は幹細胞の分化の異常に起因すると考えています。
成熟した葉状体を形成する前のゼニゴケには、背腹軸にそって並ぶ2つの幹細胞が存在します。正常な発生過程では、将来の背側に位置した幹細胞が幹細胞性を失うのに対し、腹側に位置した幹細胞は、幹細胞として機能しつづけ、周辺組織を形成していくとのモデルが過去に提唱されています(Halbsguch, 1936)。そのため、パンドン型の表現型は、背側の幹細胞も失活できないまま周辺組織を形成してしまった状態ではないかと考えています。パンドン型は、PINの制御因子に変異があるときに加え、光・重力のシグナルがない場合に起きます。そのため、光・重力のシグナルは、オーキシンとPINの働きによって、この幹細胞の運命決定のための時空間的情報をゼニゴケ組織中に生み出すのではないかと考えています。
さらに、この光・重力のシグナルによって引き起こされる幹細胞の間にできる差違は、オーキシンとPINによる体軸形成の原始的な形態である可能性が考えられます。
本現象の解明により、植物の体軸の進化においてオーキシンとPINの分子基盤が果たしてきた役割の一端が明らかになる可能性があります。そこで、光・重力のシグナルがどのような時空間的差違をゼニゴケの組織中に生み出し、幹細胞の運命決定を行うのか明らかにするための実験を行っています。
3. 環境応答時のPINクラスターとVAN3を中心とした膜ダイナミクスの動態
動物は、生育環境が自分に都合が悪くなったりすると快適な環境を求めて移動します。これに対して、植物は自分のからだのかたちを変えることによって対応します。これは、後胚発生的に器官形成を行うことができる植物だからこそできる対応方法です。
植物にとっての環境変化には、光・重力シグナルの変化、栄養状態の変化、病原菌の侵入や機械的刺激・損傷などがありますが、これらのなかでも、光・重力シグナルは、オーキシン極性輸送の制御を介して、植物の発生・成長に影響を与えます
わたしたちの研究室で発見したPINクラスターは、発生・成長過程だけでなく、光や重力、土壌環境などの様々な環境刺激に応じて状態が変化する様子が観察されています(楢本、未発表)。この外界からの刺激が、どのようにして生体内のシグナルに変換され、形態形成を制御しているのか?今後、様々な環境ストレス時のPINクラスターの動態の解析を行います。
4. 環境ストレス(土壌環境・病原菌応答)応答における細胞膜ドメイン動態
外部環境の変化に応じて、細胞膜のリン脂質の組成や分布が動的に変化することが明らかになってきました。わたしたちの研究室で解析が進んでいるVAN3は、細胞膜のリン脂質の状態によって活性・局在が制御されます(Naramoto et al., 2009)。このVAN3は、土壌環境の変化や病原菌応答においても重要な役割を果たすことを示唆する結果もあります。こちらの項目(細胞極性・体軸形成の分子機構の解明)でも述べたように、VAN3を中心とした解析により、膜環境のダイナミズムについての研究展開が可能な状況にあります。そこで今後、VAN3が実際にこれらの環境応答にどのように関わるのか、イメージングや生理学的実験を用いて明らかにします。また、VAN3が制御する最も重要な積み荷のひとつであるPIN、およびPINクラスターとの関係を明らかにすることも目指します。