植物はどのようにして、細胞から体のかたちをつくるのか?
植物の形づくりには、細胞一つひとつの「向き」が深く関わっています。細胞の中で特定のタンパク質が片側に集まることで細胞に極性が生まれ、その極性が組織の中でそろうことで、植物ホルモンの流れや成長方向、体軸や器官のパターンが決まります。
私たちの研究室では、植物ホルモンであるオーキシンの輸送を担う PIN タンパク質に注目しています。近年、私たちは PIN が細胞膜上で「PIN クラスター」と呼ばれる構造を形成することを見出しました。この PIN クラスターを手がかりとして、細胞膜上の分子配置がどのように細胞極性や植物の形づくりにつながるのかを明らかにしたいと考えています。
現在取り組んでいる主な研究課題は、以下の通りです。
1. PIN クラスターの形成と機能から探る、植物の細胞極性・体軸形成のしくみ
- PINクラスターの形成と崩壊のメカニズム
- オーキシンキャナライゼーション仮説の検証
2. 植物特有のパターン形成:葉脈パターンはどのようにして決まるのか?
3. 膜ダイナミクスに注目した細胞極性形成のしくみ
研究内容をさらに詳しく
1. PINクラスターの機能解析から探る植物の細胞極性・体軸形成のしくみの研究
PINクラスターの形成と崩壊のメカニズム
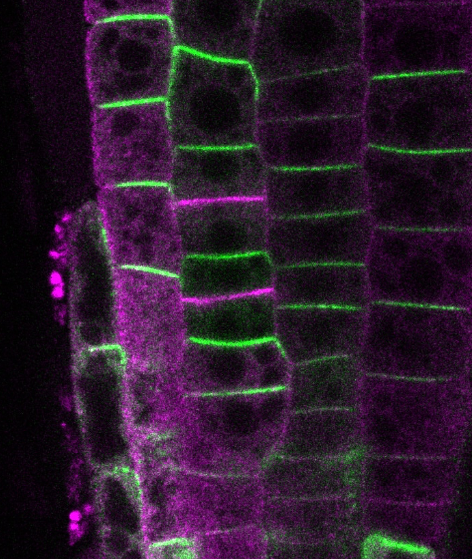
多細胞生物の発生では、細胞の極性と体軸が適切に形成されることが重要です。維管束植物では、オーキシンの極性輸送が体軸形成や器官形成に深く関わっています。この輸送方向は、オーキシン輸送体である PIN タンパク質が、細胞膜のどの側に局在するかによって決まります。
これまで PIN の極性局在については、小胞輸送によって PIN が細胞膜上をリサイクルされることで維持される、というモデルが広く考えられてきました。しかし、最近の研究から、それだけでは PIN の安定した極性局在を十分に説明できない可能性が見えてきました。私たちは、PIN が細胞膜上で形成する PIN クラスターが、PIN の局在を安定化し、細胞極性の形成に寄与する重要なしくみであると考えています。
現在、PIN クラスターの分子構造、形成メカニズム、化学的性質、そしてオーキシン輸送活性との関係を解析しています。PIN クラスターの形成と崩壊を調べることで、細胞膜上の分子集合体がどのように細胞極性をつくり、植物の体軸形成へとつながるのかを明らかにしたいと考えています。
オーキシンキャナライゼーション仮説の検証
オーキシンには、自らの流れを強めるように働く性質があると考えられており、これはオーキシンキャナライゼーション仮説として知られています。私たちは、PIN クラスターの動態、細胞内オーキシン動態の可視化、局所的なオーキシン摂動、数理モデルを組み合わせることで、オーキシンの流れと細胞極性がどのように相互作用するのかを検証していきます。
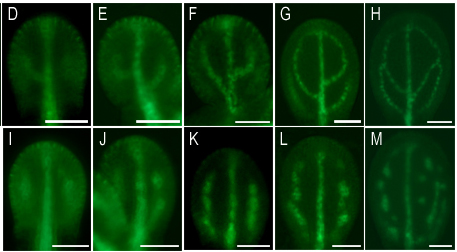
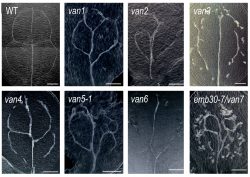
2. 植物特有のパターン形成:パターンはどのようにして決まるのか?
葉脈パターンの形成は、細胞分裂、維管束分化、細胞極性、オーキシン輸送が関わる複雑な現象です。葉脈は一見すると安定した構造に見えますが、その形成過程では、細胞レベルの分子の動きが組織全体のパターン形成へとつながっています。
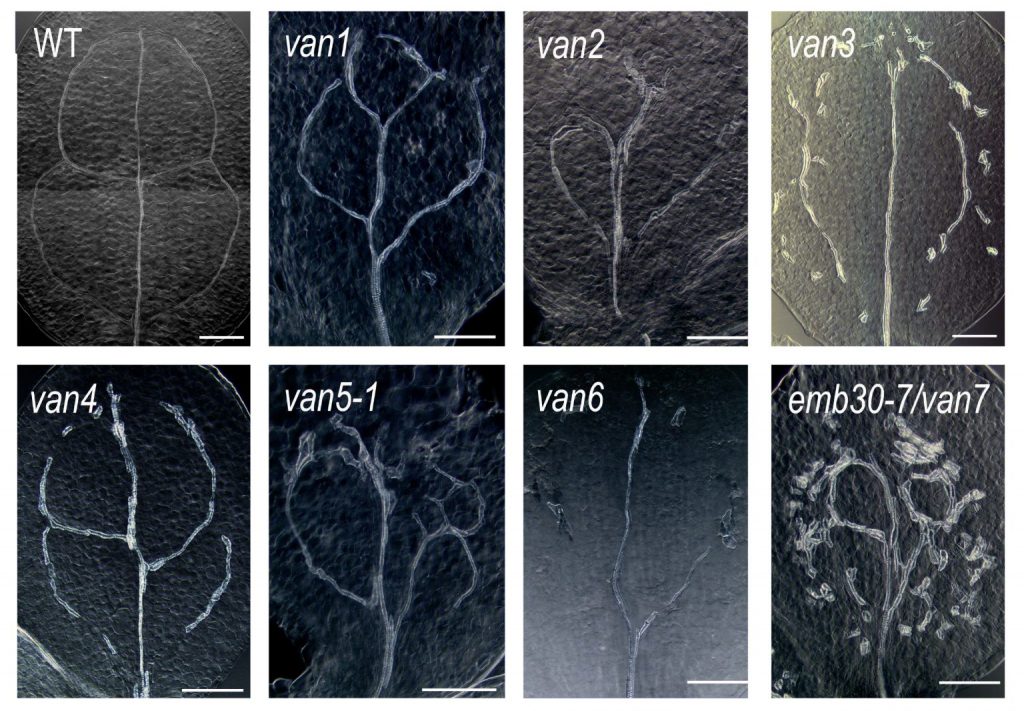
私たちはこれまでに、葉脈パターンが異常になる van 変異体を単離し、その原因遺伝子の解析を行ってきました。その結果、VAN3、VAN7、VAN4 などが小胞輸送や細胞膜環境の制御に関わり、PIN の局在制御を通じて葉脈パターン形成に影響することを見出してきました。
しかし、細胞内の分子メカニズムが、どのようにして組織全体の葉脈パターンをつくるのかは、まだ十分には分かっていません。この問題を理解するためには、分子遺伝学的解析だけでなく、ライブイメージング、オーキシン動態の可視化、細胞分裂の時空間的解析、数理モデルやシミュレーション解析を組み合わせる必要があります。
今後は、van 変異体や PIN クラスターの解析を通して、細胞内の分子のふるまいが、どのように葉脈のような組織レベルのパターンを生み出すのかを明らかにしたいと考えています。
3. 膜ダイナミクスに注目した細胞極性形成のしくみ
細胞膜は、単なる細胞の境界ではありません。細胞膜上では、リン脂質、膜タンパク質、小胞輸送、細胞骨格などが相互に関わりながら、分子が集まる場所や働く場所をつくっています。PIN の極性局在も、PIN だけで決まるのではなく、細胞膜環境や膜交通、細胞骨格との関係の中で形成されると考えられます。
私たちが研究してきた VAN3 は、細胞膜やリン脂質と関係するドメインをもつ分子であり、PIN の局在制御や葉脈パターン形成に関わります。VAN3 は細胞膜のリン脂質環境によって活性や局在が制御されることが分かっており、膜環境と細胞極性をつなぐ重要な因子であると考えられます。
今後は、VAN3 の働きを中心に、細胞膜のリン脂質環境、膜の形状、細胞骨格、小胞輸送がどのように連携して PIN の局在を制御するのかを解析していきます。また、リン脂質合成酵素 CVP2 や、VAN3 と相互作用する因子についても解析し、植物細胞における膜ダイナミクスと細胞極性形成のしくみを明らかにしたいと考えています。
さらに、VAN3 関連因子の中には、Rab や Rop などの small GTPase と関係する可能性をもつ分子もあります。small GTPase は、細胞内輸送、細胞骨格、細胞極性などに関わる重要な分子スイッチです。植物特異的な因子を介して、これらの分子スイッチがどのように協調し、細胞膜上の局所的な場を形成しているのかを明らかにすることで、植物細胞に特有の極性形成機構に迫りたいと考えています。